
Early studies of agar showed that it contained galactose, 3,6-anhydro-galactose (Hands and Peats, 1938; Percival, Somerville and Forbes, 1938) and inorganic sulfate bonded to the carbohydrate (Samec and Isajevic, 1922).
Structural studies have been based on the fractionation of agar by several methods, followed by chemical and enzymatic hydrolysis. The enzymatic hydrolysis studies of W. Yaphe have been of great importance. Subsequently the spectrochemical studies using infrared spectroscopy and nuclear magnetic resonance spectroscopy, particularly 13C n.m.r., have explained many important points in the structure of these intricate polysaccharides.
Infrared spectroscopy is the most accessible method for many laboratories. Figure 8a shows different absorption bands that have been characterized for the agar spectrum. The typical bands of a carrageenan spectrum are also shown (Figure 8b) because many of its important uses are similar to those of agar and the spectra are useful for distinguishing the two. The bands at 1 540 and at 1 640 cm-1 are especially noteworthy. They come from the proteins existing in agar and about which only a few comments have been made before. The peak at 890 cm-1 has not been identified up to the present time.
N.M.R. is of great importance when studying these structures. However the technique is difficult and it requires 13C n.m.r. equipment which only a few laboratories can afford. For this kind of work it is best to consult W. Yaphe’s papers, published from 1977 – for example, Bhattacharjee, Hamer and Yaphe, (1979); Yaphe (1984); Lahaye, Rochas and Yaphe (1986).
Agar is now considered to consist of two fractions, agarose and agaropectin. These were first separated by Araki (1937) and the results were published in Japanese so they were not readily available to some research workers. For example Jones and Peats (1942) assigned a single structure to agar defining it as a long D-galactose chain residue, joined by 1,3-glycosidic links; in the proposed structure, this chain was ended by a residue of L-galactose joined to the chain at C-4 and with C-6 semi-esterified by sulfuric acid. This false structure is still mentioned in some books on natural polymers and even in recently published encyclopedias.
AGAROSE
Interest in agarose was lost until Hjerten, working under Tiselius at the University of Uppsala, began to look for an electrically neutral polysaccharide suitable for electrophoresis and chromatography. He published an improved method of separation based on the use of quaternary ammonium salts (Hjerten, 1962). A technique for agarose preparation using polyethylene glycol was reported by Russell, Mead and Polson (1964) and later this was patented with Polson (1965) named as the inventor. Both methods gave agarose of sufficient purity to allow the study of its structure.
Figure 5 shows the type, and approximate relative quantities, of the residues that can be separated from the total hydrolysis of agarose.
Figure 6 shows agarose to be a neutral, long-chain molecule formed by b -D-galactopyranose residues connected through C-1 and C-3 with 3,6-anhydro-L-galactose residues connected through C-2 and C-4. Both residues are repeated alternately. The links between the monomers have different resistance to chemical and enzymatic hydrolysis. 1,3-a links are more easily hydrolysed by enzymes (Pseudomonas atlantica) and neoagarobiose results. 1,4-b links are more easily hydrolysed by acid catalysts and yield agarobiose units. Nevertheless 1,4-b links make the polysaccharide chain particularly compact and resistant to breakage, as is found in the peptidoglycan of bacteria. The molecular weight assigned to non-degraded agarose is approximately 120 000. This weight has been determined by sedimentation measurements and it represents 400 agarbiose (or 800 hexose) units linked together.
Figure 5 Agarose hydrolysis products
Figure 6 Agarose structure
Figure 7 Agaropectins hydrolysis products
Figure 8a Infrared spectrocopy on agar films
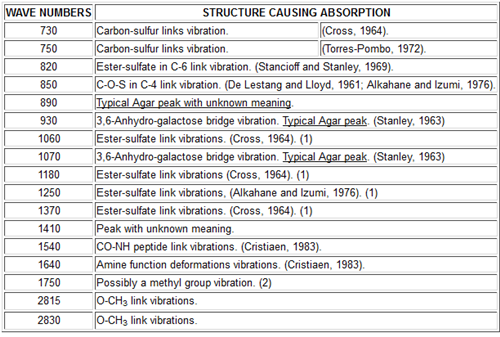
NOTES:
(1) Peaks at 1060, 1180, 1250 and 1370 are produced by sulfates but the position occupied in the chain by the sulfates is not clearly seen in Agar due its low content of sulfates (< 2%).
(2) Peak at 1750 not attributed up to this moment could be caused by methyl groups as Agar with 6-methyl forms a peak at 1780 cm-1.
Figure 8b Infrared spectrocopy on carrageenan films

NOTES. –
1.- A peak at 831 CM-1 wide is mentioned in the Bibliography to correspond to a 3-Sulfates mixture.
-O-SO equatorial vibration on C-2 of a galactose linked in (1 ® 3) ring.
-O-SO vibration on C-2 of a galactose linked in (1 ® 4) ring.
-O-SO vibration on C-6 of a galactose ring.
2.- Carrageenans have wide and strong absorption bands in 1,000-1,100 CM-1 region which are typical in all polysacharides.
3.- Maximum absorption is given by 1,065-1.020 CM-1 for all carrageenan types (Kappa, Iota, Lambda, etc.)
This clarifies the information in Figure 6. However it should be noted that, depending on the origin of the raw material, some units of 3,6-anhydro-L-galactose are replaced by L-galactose. Also some D-galactose and L-galactose units can be methylated and it is said they can be in fact 6-0-methyl-D-galactose and 2-0-methyl-Lgalactose. This methylation, arising from the seaweed used in the process, determines the agarose gel point and therefore that of the agar it comes from. D-xylose has been found in very small quantities from hydrolysed agarose but it has not been possible to assign it a position in the structure.
Polar residues such as pyruvic and sulfuric acids are also found in small quantities. They may come from the small amounts of agaropectin lef in the agarose after its preparation but in our opinion sulfate and pyruvate groups remain linked in small quantities to the agarose structure, depending on the seaweed used in agar production. We follow the traditional definition of agaroses as those products obtained as the non-charged fraction after using a classical separation technique such as the precipitation with quaternary ammonium salts by Hjerten. On the other hand, in spite of the copious bibliography on this matter (we have seen 14 different basic methods to prepare agarose), none of the methods permits an agarose preparation free of electronegative charges. Many researchers have used two or three fractionating methods successively, in order to improve the separation and reduce the amount of electronegative groups present. In spite of all these efforts, these groups could not be eliminated. To cancel the electroendosmotic flow, which might be induced by these electronegative groups, it has been necessary to fix electropositive groups or use some other means so as to reduce the migration of cations (and their solvation water molecules) fixed to electonegative groups. Consequently we consider the agarose theoretical structure a chimerical dream to which we get closer each time by using more refined fractionation methods although perhaps, in practice, it may not exist at all in agar and the agarophyte seaweeds.
Nowadays commercial agaroses for use in biochemical separation techniques have to be chemically modified, so that their structure is different from the agarose as it is extracted from the seaweed, Phycologists should be aware that this is so, unless the manufacturer states that the original chemical structure has not been modified.
Agarose is responsible for the gelling power as we know it in agar. This is a gelation in aqueous media with a very small reactivity with cations and proteins and this differentiates agar from carrageenan.
AGAROPECTIN
Agaropectin (or better, the agaropectins) have a low gelling power in water. At the present time, a specific structure has not been assigned to the agaropectins. It is customary to say that they are formed by alternating units of D-galactose and L-galactose, and that they contain all the polar groups existing in agar.
Figure 7 shows the residues obtained by hydrolysis; among them, sulfated and pyruvate residues are evident. It has been verified that L-galactose 6-sulfate and D-galactose 4-sulfate are the major sulfate residues in agar. From small to moderate quantities of 3,6-anhydro-L-galactose have also been detected. These small quantities vary depending on the origin of the seaweed, on the harvesting season, on the treatment applied during the agar manufacturing process and on the treatment used during the agarose separation process.
The presence of 4,6-0-(1-carboxyethylidene)-D-galactose has also been verified, making the position of pyruvic acid in the structure perfectly clear. This unit is relatively important in agaropectin but in agarose it appears in much lower levels, as mentioned previously, probably because agarose has terminal units of 4,6-0-(1-carboxyethylidene)-D-galactose. The quantity of pyruvic acid in agar and agarose varies widely depending on the seaweeds used as raw material; we have verified quantities between 0.2-2.50% in agar and 0.02-1.30% in agarose. In this regard the work of Hirase (1957) is very interesting and explanatory.
These variations, that sometimes can be very important, appear even in seaweeds of the same class harvested a short distance from each other and seem to be permanent and depend on the growing locations. Over a period of several years (more than 10 in some cases) we have studied different Gelidium or Gracilaria harvesting areas in Europe, Asia and America, verifying the persistence of this phenomenon that can be caused by microclimatic differences. In our opinion the differences in cations existing in certain habitats also can be a cause. Naturally the different types and species cause differences that are very important sometimes in the agarose and agaropectin structures.
In Figure 7, D-galactose 2,6-disulfate has been included because we think we have identified it in small quantities in the agaropectins of some seaweeds grown in difficult conditions (“El Niño” phenomenon). These agaropectins had high viscosity, that was also apparent in the agar from which they came, along with a lower gelling power. In cases where this sulfated residue is found, the agaropectin and the agar have undesirable properties. Also shown in Figure 7 are D-galactose and L-galactose which appear to be modular units of agaropectin. Glucuronic acid is present only in traces (like the D-xylose found in agarose).
So while the basic structure of agaropectin consists of alternating D-galactose and L-galactose, D-galactose can be substituted by D-galactose 4-sulfate, by 4,6-0-(1-carboxyethylidene)-D-galactose in certain terminal chain positions or even possibly by D-galactose 2,6-disulfate, while part of L-galactose can be replaced by 3,6-anhydro-L-galactose. These different substitutions of the basic monosaccharide give an enormous number of possible structures.
McCandless used an immunochemical method to detect different carrageenan fractions with great sensitivity (Di Ninno and McCandless, 1978 and 1978a). A similar method might be applied to studying the different kinds of agaropectins in regard to their different seaweed origins, as well as the posible evolution of the structure of agaropectins during the life of the seaweed. To do this it is necessary to take into consideration the different fractions preceding the series of biochemical transformations produced by the algal enzymatic mechanisms which result in certain terminal fractions (one of which may be agarose). The current possibilities through monoclonal antibodies would allow an improvement of the sensitivity and selectivity of the method used by McCandless.
Source: FAO
